Эмбриогенез насекомых. Эмбриональное и постэмбриональное развитие насекомых
Развитие насекомых делится на два периода: зародышевое, или эмбриональное (внутри яйца), и внеяйцевое, или постэмбриональное. Развитие зародыша в яйце начинается дроблением ядра с момента оплодотворения. Яйцо имеет круглую, овальную, и др. форму и представляет собой одну крупную клетку, включающую питательный желток, необходимый для роста и развития зародыша. Снаружи яйцо покрыто оболочкой (хорион), играющей роль скорлупы. На одном из полюсов яйца имеется одно или несколько мельчайших отверстий (микропиле), через которые проникают сперматозоиды при оплодотворении. Яйцо после откладки часто всасывает влагу из окружающей среды и вследствие этого увеличивается в 2-3 раза.
Зрелые яйца самки откладывают поодиночке или группами на листья, ветви и стволы деревьев, на почву, травянистую растительность и другие предметы. Часто яйца бывают погружены в субстрат (в древесную ткань или почву) или защищены пушком, снятым с конца -брюшка самки (у златогузки, непарного шелкопряда и других бабочек), или щитком, образовавшимся из выделений придаточных желез (у зеленой узкотелой златки).
Развитие зародыша в яйце продолжается от нескольких дней до месяца и более. Оно заключается в ряде сложных преобразований и заканчивается вылуплением личинки, прогрызающей оболочку яйца
Строение яйца
Яйцо насекомого представляет собой крупную клетку, покрытую снаружи хорионом, обычно имеющим характер скорлупы, под которой находится истинная, или желточная, оболочка. Хорион образован из выделений эпителиальных клеток яйцевой трубки яичника, и на его поверхности часто можно обнаружить более или менее сильно выраженные скульптурные образования, представляющие отпечатки клеток фолликулярного эпителия. На поверхности хориона имеется отверстие - микропиле, которое служит для проникновения сперматозоида.
Типы яиц и характер их откладки. Кроме размера, яйца могут различаться формой, скульптурой хориона, окраской. По форме они чаще бывают овальными, удлиненно-овальными, шаровидными. Вместе с тем в ряде случаев форма яиц очень типична для нескольких видов одного рода, а нередко и семейства. Так, очень характерны бутылковидные яйца у капустной и репной белянок, бочонковидные - у клопов щитников, полу шаровидные - у совок и т. д.
Наружная поверхность яиц, т.е. скульптура хориона, чаще гладкая, но может иметь продольные или поперечные ребрышки, морщинки, бороздки и т. д. Так, у яиц бабочек совок хорион имеет радиальные ребрышки, у яиц капустной и репной белянок - крупные продольные и мелкие поперечные ребрышки, у яиц некоторых кокцинеллид - мелкоячеистую скульптуру и т. д.
Самки насекомых откладывают большое количество яиц в один или несколько приемов. При этом размещают их по одному, небольшими группами или кладками по 100, 200 и более яиц, что типично для данного вида. Так, капустная белянка откладывает яйца на листья растений кладками до 200 яиц в каждой, а репная белянка - по одному.
Кладки яиц могут быть отложены открыто на растениях, например у капустной белянки, кольчатого шелкопряда, кокцинеллид, или скрыты в субстрате (почва, ткани растений), прикрыты волосками с брюшка самки или снаружи защищены стенками кожистой капсулы. Так, кладки яиц непарного шелкопряда и златогузки настолько плотно прикрыты волосками с брюшка самки, что сверху совершенно не видны, а кладки яиц тараканов и богомолов находятся внутри кожистой капсулы - оотеки, образованной застывшими выделениями придаточных желез самки. Очень своеобразны также кубышки саранчовых, содержащие от 5-15 яиц у атбасарки до 140 яиц у пустынной саранчи. Секрет придаточных желез самки не только обволакивает их, но и цементирует прилегающие частицы почвы.
Развитие зародыша
Эмбриональное развитие начинается с дробления ядра и передвижения образовавшихся дочерних ядер с небольшими участками протоплазмы к периферии яйца. Здесь они образуют сплошной слой клеток - бластодерму, охватывающую скопление желтка. В дальнейшем бластодерма дифференцируется на зародышевую и внезародышевую зоны. Клетки зародышевой зоны делятся более интенсивно и образуют с брюшной стороны яйца зародышевую полоску.
В процессе дальнейшей дифференциации зародышевой полоски образуется ее наружный листок - эктодерма, по краям формирующая двухслойную складку, а сама полоска несколько погружается в желток. Нарастающие края складки смыкаются друг с другом и срастаются, в результате чего зародышевая полоска покрывается двумя слоями клеток - зародышевыми оболочками. Наружную оболочку называют серозой, внутреннюю - амнионом. Между амнионом и развивающимся зародышем образуется полость, заполненная жидкостью, имеющей защитное и пищевое значение для зародыша. Наличие оболочек объясняет повышенную устойчивость яиц насекомых после прохождения этого этапа эмбриогенеза к инсектицидам, а также к иссушению.
По мере образования зародышевых листков начинает формироваться зародыш насекомого. Из эктодермы образуются кожные покровы тела; путем впячивания эктодермы с обоих полюсов возникают ротовое и анальное отверстия, передний и задний отделы кишечника, зачатки будущих трахей и нервной системы. Из внутреннего зародышевого листка - энтодермы образуется эпителий средней кишки, из среднего - мезодермы формируются мышцы, жировое тело, спинной сосуд, оболочка половых желез.
Сформировавшийся зародыш, превратившийся, по существу, в личинку, заполняет все яйцо, начинает интенсивно двигаться, набирает воздух в трахеи, заглатывает амниотическую жидкость, увеличиваясь в объеме, освобождается от зародышевых оболочек, разрывает хорион и вылупляется, т. е. выходит наружу.
Обычно эмбриональное развитие насекомых длится от 2 до 10 дней, реже 2-3 недели, в зависимости главным образом от температуры среды. При длительной остановке в развитии - эмбриональной диапаузе - продолжительность фазы яйца у некоторых насекомых (саранчовые, мухи цветочницы и др.) может достигать 6-9 месяцев.
В середине, а скорее даже в первой половине прошлого века многие области науки и техники пережили небывалый расцвет, поднявшись на волне всеобщей веры, что развитие фундаментальной науки может быстро реализоваться, как это было, например, с атомной бомбой или с вакцинацией. Были открыты генетический код и структура ДНК, появились новые науки, взявшиеся изучать исконные вопросы медицины, физиологии и систематики с молекулярной точки зрения. Под этим углом была «пересмотрена» и биология развития (и, в частности, эволюция развития), пытавшаяся выяснить: каким образом гены управляют эмбриогенезом, какова динамика аллелей в популяции, и, наконец, как на уровне ДНК возникают новые признаки?
Ответы на эти вопросы важны не только с теоретической точки зрения. Устранение физического неравенства и боли, продление жизни и борьба с болезнями... Наша цивилизация вплотную подошла к решению этих задач генетическим способом: исправление плохих копий генов и встаивание чужеродного генетического материала, способствующего борьбе с болезнями. Один из примеров – трансгенные растения, несущие гены инсектицидов и устойчивости к заболеваниям. Другой и совсем недавний – создание гипоаллергенной кошки путем модификации генов, кодирующих аллергенную компоненту шерсти.
Конечно, применение новейших технологий не всегла приводит к успеху, и, исправляя одно, мы поневоле разрушаем другое. Так произошло, например, с неизлечимой болезнью SCID-X1, вызываемой мутацией гена, ответственного за созревание определенного типа лимфоцитов. Лекарственная терапия в данном случае неэффективна. Зато генно-терапевтический способ оказался вполне удачным. У пациента брали клетки-предшественники и с помощью ретровируса встраивали в их ДНК правильную копию гена. «Исправленные» клетки вводили больному, что приводило к восстановлению его иммунной системы. Однако один больной, у которого иммунная система восстановилась, умер от рака крови. Оказалось, что ретровирус встроился в антираковый ген и разрушил его.
Каким образом вносить сбалансированные изменения в генотип? Почему природа может, а мы – нет? Как создавать новые признаки по своему желанию? В конце концов, почему мухи разносят болезни, а сами не болеют?
Спонсор публикации статьи – крупнейший поставщик массажной продукции, официальный представитель Yamaguchi, US Medica, Fujiiryoki – компания «Ямагучи», предлагающая массажные столы, кресла, накидки и прочие массажеры. Матрас массажный Ocean, так же поставляемый компанией в Россию, является интересным приобретением для домашнего использования. С помощью 26 воздушных подушек матрас проводит массаж тела лежащего на нем человека, сначала расслабляя его, потом плавно разрабатывая и потягивая позвоночник, что имитирует сеанс мануальной терапии. Использование массажера позволяет избавиться от боли в спине и пояснице, увеличивает подвижность и свободу движений.
Генетика и семантика
Вопросы, конечно, интересные, но есть и более важные аспекты. Вот, например, семантическое значение гена: у каждого ли гена есть строго определенная роль? Влияет ли на ген его окружение? Где и как записано в ДНК, что у одного человека нос длиннее, а у другого короче; у двукрылых насекомых два крыла, а у бабочек – четыре? Есть ли унифицированные законы эволюции генов? Можем ли мы использовать их в создании новых генов по нашему желанию?
Как уже говорилось, эволюция развития изучает изменение генов по мере возникновения новых признаков. Изучение ведется в двух направлениях.
Первое направление ориентируется на признаки, детально изученные с молекулярной и физиологической точек зрения, проводя реконструкцию изменений гена во времени, вызвавших возникновение новых признаков, а также устанавливая факторы, закрепляющие эти модификации. Наиболее удобная модель для данной области – нейтральная селекция, когда признак не меняется вообще, а гены претерпевают взаимокомпенсирующие модификации.
Для второго направления отправной точкой служит хорошо изученное событие развития, по-разному выраженное у двух организмов. Детальное знание генетики процесса в одном организме облегчает идентификацию гена, отвечающего за изучаемый признак у другого. В этой статье мы рассмотрим в основном второе направление.
Решение подобных задач предшествует внесению в человеческий или иной организм новых «улучшающих» признаков по нашему усмотрению. Для успешного их осуществления должны быть выполнены следующие условия.
1. Необходимы два или более модельных организма для установления сравнительной системы.
2. Изучаемые признаки модельных организмов должны быть легко различимы.
3. Механизмы или физиология развития исследуемого признака в обоих организмах должны быть точно установлены.
4. Геномная ДНК обоих организмов должна быть отсеквенирована, т.е. известна ее полная последовательность.
5. Организмы не должны отстоять слишком далеко друг от друга на эволюционном древе. Наша задача – найти наименьший генетический комплемент, соответствующий возникновению нового признака или фенотипической разнице, поэтому нас интересуют организмы, максимально близкие по ДНК, но анатомически разные.
Найти подобные два организма непросто. Во-первых, исторически эмбриология стремилась выработать унифицированный язык, подходящий для описания развития в любом организме, и вследствие этого фенотипически разный эмбриогенез зачастую описывается одинаково для филогенетически близких животных. Во-вторых, возможность секвенировать целые геномы появилась лишь недавно и даже теперь количество «отсеквенированных» организмов ограниченно.
Тем не менее, на сегодняшний день существует согласованный круг молекулярно-эволюционных задач и определены типы сравнительных систем, подходящих для их решения. Так, например, для изучения нейтральной селекции и выделения консервативных, а следовательно, и функционально важных элементов ДНК используется выборка родственных видов из насекомых семейства Drosophilidae . Раннее развитие мух данного семейства практически не содержит различий, по крайней мере – в рамках некоторых процессов, генетика которых хорошо изучена. Подобный же анализ применяется к «секвенированным» позвоночным животным (человек, крыса, мышь, рыбка-зебра). Он позволяет идентифицировать законсервированные элементы ДНК, повторяющиеся у этих организмов. Существует мнение, что подобная консервация свидетельствует о функциональной значимости фрагментов ДНК.
Я в своей работе использую сравнительный анализ развития зародышей плодовой мухи Drosophila melanogaster и малярийного комара Anopheles gam biae . Меня интересует следующий вопрос: каким образом изменение морфологии в развитии животных отражено в ДНК? Можно ли превратить одно животное в другое путем нескольких генетических модификаций? Каковы законы эволюции генетических сетей, ответственных за формообразование?
Филогенетический метод и реконструкция истории развития вида
Эволюционный взгляд на биологическое формообразование предполагает, что каждый момент истории был представлен в природе определенным разнообразием видов, часть из которых вымирала, а часть служила предшественниками новых видов. Однако достоверно мы можем судить лишь о видовом многообразии, существующем на данный момент. Все остальное – область более или менее достоверных гипотез.
Как же реконструировать историю возникновения нового вида (признака или органа), не имея исторического материала? Для этого используется филогенетический метод: история возникновения вида реконструируется по присутствию в современной фауне признаков, присущих этому виду.
Представим себе лежащие на столе пустую тетрадь, исписанную тетрадь, толстую книгу, глянцевый журнал, ежедневник и электронную записную книжку. Все эти предметы, изготовленные в наше время, служат для передачи, как правило, словесной информации. Большинство из них сделаны из бумаги и вследствие этого имеют одинаковую структуру.
Исходя из преобладания бумажных носителей над электронными, можно предположить, что электронная записная книжка – сравнительно недавнее изобретение. Остальные вещи различаются количеством страниц, но тем не менее состоят из них. Из этого можно сделать вывод, что когда-то предшественником этих изделий была книга всего из одной страницы, а переплет – позднейшее изобретение. Рассуждая подобным образом, мы можем прийти к выводу, что eще более древним предшественником этих изделий была целлюлоза, т.е. древесина.
В этой статье мы рассмотрим эмбриогенез комара и мухи и, в частности, развитие так называемых экстраэмбриональных мембран. У комара их две (амнион и сероза), а у мухи одна (амниосероза). Практически у всех насекомых, кроме небольшого подотряда двукрылых, две мембраны, как у комара. Это позволяет нам предположить, что предшественник современных мух имел две экстраэмбриональные мембраны и комар является моделью этого предшественника.
Как работает ген?
Несмотря на то что основным результатом экспрессии гена служит продукция белка, лишь малая часть гена участвует в его кодировании. Этот участок находится внутри зоны, покрытой непрерывной последовательностью чередующихся экзонов и интронов. Интроны удаляются из конечной «кодирующей РНК», а экзоны сшиваются вместе.
Значительная часть гена не транскрибируется в РНК, а содержит компактные зоны, содержащие участки (сайты) связывания транскрипционных факторов (обычно это белки). Эти зоны называются энхансерами. Чем лучше сайты в энхансере, тем сильнее связывание транскрипционного фактора, тем более чувствителен ген к низким концентрациям активаторов (транскрипционных факторов, усиливающих продукцию РНК) или репрессоров (белков, блокирующих продукцию РНК).
На рисунке 1 показан ранний эмбрион мухи, покрашенный специальным образом для детекции РНК гена eve (рис. 1, а ). Вторая полоса, собственно eve , выделяется дополнительно с помощью синего красителя. Ниже приведена диаграмма гена eve , в которой начало транскрибируемой области eve показано стрелкой, а конец – нуклеотидной последовательностью конца последнего экзона (рис. 1, б ). Регуляторные участки (энхансеры) обозначены подписанными четырехугольниками. Цифры в них обозначают номера полос, регулируемых данным энхансером.
Под диаграммой гена размещена диаграмма регуляторного участка, управляющего экспрессией второй полосы (рис. 1, в ), затем небольшой фрагмент этого участка с нуклеотидами, вовлеченными в связывание конкретных транскрипционных факторов (рис. 1, г ).
Чем ближе родство двух организмов, тем меньше отличий в последовательности ДНК конкретного гена в обоих геномах. Копии одного гена в разных геномах называются ортологами (например, мышиная ДНК-полимераза является ортологом человеческой). Исследования показали, что некодирующие участки ортологичных генов содержат больше отличий, чем кодирующие. Необходимость «удерживать» последовательность белка от мутаций препятствует их накоплению в кодирующих областях. Таким образом, некодирующие участки гена (содержащие энхансеры и другие регуляторные элементы) значительно более пластичны с эволюционной точки зрения.
На данный момент преобладает мнение, что формообразование и модификация признаков в первую очередь происходят за счет разницы в некодирующей ДНК.
Эмбриогенез насекомых. Генетические сети
Удобную эмбриологическую модель представляют собой яйца насекомых. Во-первых, в силу сравнительной легкости поддержания колонии насекомых и сбора яиц, во-вторых, в связи с относительно быстрыми темпами развития: у плодовых мух – 24 ч от оплодотворения ооцита до вылупления личинки. Небольшие размеры облегчают изготовление срезов и зачастую позволяют умещать целый эмбрион в поле зрения микроскопа.
Создание описательной базы по раннему развитию насекомых шло по мере развития методов микроскопии. С первой половины XX в. применение физиологических методов (пережимание яиц, трансплантация цитоплазмы из разных частей ооцита, трансплантация фрагментов эмбриона) позволило установить наличие эмбриональных центров, производящих растворимые факторы, определяющие судьбу клеток в зоне их действия. Однако истинным прорывом стало применение технологии мутационного скрининга (отработанной на бактериях и дрожжах в 1960–1970 гг.) к изучению генетики развития плодовых мух Drosophila melanogaster .
 К моменту проведения этих экспериментов было понятно, что гены являются фундаментальной единицей биохимических процессов, ответственной за отдельный признак в наиболее примитивных случаях. Однако считалось что глобальные признаки (определение эмбриональных осей развития: антеро-каудальной (между головой и хвостом) и дорсо-вентральной (т.е. спинно-брюшной); детерминация типа и числа сегментов; пространственная спецификация конечностей и т.д.) являются полигенными и едва ли будут расшифрованы в ближайшие десятилетия. К удивлению исследователей, в первых же скринингах были получены мутантные мухи, производящие эмбрионы, целиком вентрализованные, дорсализованные, потерявшие строго определенный сегмент или группу конечностей.
К моменту проведения этих экспериментов было понятно, что гены являются фундаментальной единицей биохимических процессов, ответственной за отдельный признак в наиболее примитивных случаях. Однако считалось что глобальные признаки (определение эмбриональных осей развития: антеро-каудальной (между головой и хвостом) и дорсо-вентральной (т.е. спинно-брюшной); детерминация типа и числа сегментов; пространственная спецификация конечностей и т.д.) являются полигенными и едва ли будут расшифрованы в ближайшие десятилетия. К удивлению исследователей, в первых же скринингах были получены мутантные мухи, производящие эмбрионы, целиком вентрализованные, дорсализованные, потерявшие строго определенный сегмент или группу конечностей.
На рисунке 2, а показаны некоторые последовательные стадии эмбриогенеза плодовой мухи. На всех иллюстрациях в этой статье эмбрионы показаны так, что головной отдел находится слева, а хвостовой – справа. Фотографии сделаны с помощью сканирующей электронной микроскопии. На рисунке 2, б – эмбрион мухи, мутантной по гену bcd . Заметно, что головной отдел (слева) копирует морфологию хвостового отдела. Таким образом, были идентифицированы несколько иерархических групп взаимосвязанных генов, ответственных за раннюю пространственную разбивку эмбриона.
По большей части эти гены кодируют транскрипционные факторы. Внутри группы гены нижнего уровня являются мишенью для активации или ингибирования генами, стоящими выше по иерархии. Такая структура групп напоминает компьютерную или нейронную сеть, и далее мы будем называть ее генной сетью.
 Рассмотрим вкратце генную сеть, управляющую разбивкой тела в направлении голова–хвост (рис. 3). У дрозофилы исходными элементами этой сети являются гены bcd
и nos
. РНК этих генов размещается соответственно в головном и хвостовом отделах яйца. РНК другого важного гена cad
равномерно распределена по яйцу. Белковый продукт, синтезирующийся с РНК гена bcd
(транскрипционный фактор BCD), распределен по раннему эмбриону в виде градиента от головы к хвосту.
Рассмотрим вкратце генную сеть, управляющую разбивкой тела в направлении голова–хвост (рис. 3). У дрозофилы исходными элементами этой сети являются гены bcd
и nos
. РНК этих генов размещается соответственно в головном и хвостовом отделах яйца. РНК другого важного гена cad
равномерно распределена по яйцу. Белковый продукт, синтезирующийся с РНК гена bcd
(транскрипционный фактор BCD), распределен по раннему эмбриону в виде градиента от головы к хвосту.
Транскрипционный фактор BCD способен ингибировать трансляцию белка CAD с РНК гена cad , создавая противоположный BCD градиент CAD, исходящий из хвостового отдела яйца.
В головном отделе эмбриона BCD активирует экспрессию гена hb , которая убывает по мере истощения количества BCD в градиенте, а также экспрессию гена gt , которая убывает быстрее, чем hb , потому что регуляторная ДНК gt менее чувствительна к низким концентрациям bcd , чем энхансер hb .
В заднем отделе эмбриона hb активируется градиентом CAD. Gt тоже экспрессируется в хвостовом отделе, причем его зона экспрессии находится впереди зоны hb .
Ген Kr активируется BCD и одновременно ингибируется HB. Регуляторные последовательности этого гена более чувствительны к низким концентрациям BCD, нежели энхансер hb , поэтому область его экспрессии простирается далее области, занятой HB. Вследствие ингибирования в головном отделе создается характерная колоколообразная зона экспрессии гена Kr . Аналогичные механизмы с CAD активатором вместо BCD вовлечены в образование колоколообразного домена экспрессии гена kni .
Гены hb , Kr , kni в совокупности называются GAP-генами (от англ. gap – пробел), поскольку в случае мутации личинка мухи теряет сегменты, расположенные в зоне нормальной экспрессии GAP-гена.
Сегментация эмбриона является кульминационной ступенью в раннем развитии насекомых. На генетическом уровне повторяющиеся сегменты возникают вследствие повторной экспрессии определенных генов вдоль оси эмбриона голова–хвост. Примером может служить начальная экспрессия мушиного гена eve в каждом четном сегменте зародыша.
Экспрессия генов сегментации, таких как eve , непосредственно регулируется GAP-генами. Более того, фактически каждой полоске экспрессии соответствует свой регуляторный элемент, независимо позиционирующий этот домен в эмбрионе.
Похожие и разные стадии в развитии плодовых мух и малярийного комара
 На первый взгляд, личинки комара и мухи сильно различаются (рис. 4). Это неудивительно: на стадии личинки комар – водное насекомое, обладающее особыми органами дыхания и специальным челюстным аппаратом. Личинка плодовой мухи обитает в гниющих фруктах. Ее строение (короткие щетинки на кутикулярной оболочке) оптимально для облегчения ввинчивания в мякоть плодов. Однако с эмбриологической точки зрения зародыши обоих насекомых практически неразличимы – они содержат одинаковый набор органов и проходят через идентичные ступени развития.
На первый взгляд, личинки комара и мухи сильно различаются (рис. 4). Это неудивительно: на стадии личинки комар – водное насекомое, обладающее особыми органами дыхания и специальным челюстным аппаратом. Личинка плодовой мухи обитает в гниющих фруктах. Ее строение (короткие щетинки на кутикулярной оболочке) оптимально для облегчения ввинчивания в мякоть плодов. Однако с эмбриологической точки зрения зародыши обоих насекомых практически неразличимы – они содержат одинаковый набор органов и проходят через идентичные ступени развития.
На самых ранних стадиях эмбриогенеза оплодотворенное яйцо представляет собой одну гигантскую клетку, заполненную питательным желтком. Первые 14 ядерных делений происходят без дробления ооцита. Ядра выстраиваются по периферии клетки, образуя так называемый ядерный бластодерм. После 14-го деления клеточные мембраны стремительно прорастают между ядрами, разделяя их на клетки. Эта стадия называется клеточный бластодерм.
У насекомых отряда двукрылых (Diptera ) на стадии клеточного бластодерма клеточная спецификация практически завершается: территориально представлены фактически все будущие органы личинки. То есть можно предсказать, предшественником или частью какого будущего органа является та или иная клетка.
Следующая стадия характеризуется морфологическим обособлением так называемой эмбриональной пластинки. Клеточная бластодерма сдвигается в вентральном (брюшном) направлении, таким образом вентральные клетки (предшественники мезодермы, образующей внутренние органы) приобретают колоннообразную форму (вытянутую по вертикальной оси), а дорсальные клетки уплощаются.
Интересно, что дорсальные (спинные) клетки и их потомки не будут представлены в личинке. Дорсальные клетки образуют так называемые экстраэмбриональные мембраны, клетки которых отмирают на завершающих стадиях эмбриогенеза. Напротив, латерально-вентральные (боковые и брюшные) клетки являются предшественниками всех органов личинки.
Таким образом, континуум клеток-предшественников личинки не замкнут с дорсальной стороны, что и позволяет говорить о так называемой эмбриональной пластинке.
На следующей стадии начинается рост – вытяжение эмбриональной пластинки. До сих пор развитие эмбрионов комара и мухи происходило одинаково, но на этой стадии наиболее наглядно проявится разница.
В случае мушиного зародыша эмбриональная пластинка растет, огибая задний полюс яйца. По мере роста она сминает дорсальные клетки. В случае комара эмбриональная пластинка не сминает дорсальные клетки, но, напротив, проползает под ними. Таким образом создается зачаток трехслойной структуры.
Нижний слой – эмбриональная пластинка, промежуточный слой экстраэмбриональных клеток, который называется амнионом, и внешний (верхний) – сероза. Амнион начинает мигрировать по поверхности эмбриональной пластинки и растягивает вокруг нее серозу. В итоге эмбрион комара, как и практически всех остальных насекомых, кроме «высших двукрылых», оказывается внутри серозного мешка.
Такая структура очень важна для развития насекомых, особенно тех, чьи яйца должны переживать период засухи (яйца многих видов комаров устойчивы к высыханию). Дополнительная хитинсодержащая оболочка, секретируемая серозой, предохраняет эмбрион от высыхания. Мушиные эмбрионы не используют этот механизм.
Различия генетических сетей, управляющих антеро-каудальной разметкой
Таким образом, с анатомической точки зрения развитие мушиного и комариного эмбриона по антеро-каудальной оси происходит одинаково, однако в процессах, происходящих вдоль дорсовентральной оси, наблюдается существенная разница. Есть ли отличия в структуре генетических сетей, управляющих пространственной разбивкой эмбриона по этим осям?
Для ответа на этот вопрос мы решили исследовать экспрессию генов, ответственных за эмбриогенез в зародышах мухи и комара. Для локализации экспрессии использовали химически меченые антикомплементарные пробы к РНК интересующих нас генов.
Ген-регулятор сегментации eve выглядит одинаково в мушином и комарином зародыше, что соответствует филотипичности этой стадии у насекомых и членистоногих в целом. GAP-гены Kr и kni , регулирующие независимую экспрессию разных полосок eve , тоже занимают сравнительно похожие домены. Расположение головных доменов hb и gt также одинаково. Однако в то время как у мухи задняя полоска hb расположена позади каудального домена gt , у комара все наоборот: каудальный домен gt расположен позади каудального домена hb .
Эта казалось бы незначительная разница имела бы катастрофические последствия для развития комара, если бы регуляция экспрессии eve осуществлялась так же, как у мухи. Дело в том, что шестая и седьмая полоски eve у мухи расположены между зонами экспрессии репрессоров kni и hb . У комара задние полоски kni и hb (генов, ингибирующих экспрессию eve) оказываются столь близко друг к другу, что поместить даже одну полоску между ними невозможно. По всей видимости, экспрессия шестой и седьмой полосок eve у комара регулируется комбинацией kni и gt .
Как уже было замечено выше, у мухи третья и седьмая полоски регулируются одновременно одним энхансером. Аналогично четвертая и шестая полоски тоже регулируются совместно одним регуляторным элементом. Судя по одинаковому расположению доменов экспрессии GAP-генов в передней части зародыша, регуляция третьей и четвертой полосок eve у мухи и комара не различается. Однако механизмы установления шестой и седьмой полосок различны. Следовательно, у комара есть дополнительные независимые регуляторные элементы (энхансеры), регулирующие шестую и седьмую полоски. То есть у комара в два раза больше энхансеров eve , чем у мухи.
Плодовая муха – значительно более динамичный организм, чем комар. Мухи откладывают больше яиц и развиваются значительно быстрее. Очевидно, что редукция и оптимизация регуляторных элементов у мухи суть проявления более динамичного жизненного цикла.
В данном случае различия в архитектуре генетической сети регуляции гена eve не приводят к изменению картины распределения продукта гена в эмбрионе, т.е. осуществляется компенсаторная, нейтральная селекция гена, не приводящая к изменениям в морфологии филотипической стадии.
Возникает вопрос: можно ли предсказать альтернативные формы генетических сетей исходя из знания одного из вариантов? Как происходит компенсаторная эволюция в историческом плане?
Различия генетических сетей, управляющих дорсо-вентральной разметкой
 Противоположный сценарий реализуется вдоль дорсо-вентральной оси. Основой данного элемента разбивки является внутриядерный градиент транскрипционного фактора DL, который в зависимости от контекста может быть как активатором, так и репрессором (рис. 5). DL aктивирует серию генов вдоль дорсо-вентральной оси. Чем более чувствителен энхансер гена к белку DL, тем выше расположена зона его экспрессии.
Противоположный сценарий реализуется вдоль дорсо-вентральной оси. Основой данного элемента разбивки является внутриядерный градиент транскрипционного фактора DL, который в зависимости от контекста может быть как активатором, так и репрессором (рис. 5). DL aктивирует серию генов вдоль дорсо-вентральной оси. Чем более чувствителен энхансер гена к белку DL, тем выше расположена зона его экспрессии.
Другим кардинальным элементом данной генетической сети является ген dpp . Он кодирует внеклеточный белок, экспрессия которого ингибируется DL. DPP диффундирует в вентральном направлении и включает в клетках транскрипционный фактор MAD (pMAD – в активной форме), который активирует гены в дорсальной части зародыша.
Мушиный ген sog – один из наиболее чувствительных к DL генов (его энхансер содержит очень сильные сайты связывания DL). Зона экспрессии sog расположена в латеральной части эмбриона мухи. В вентральных клетках мушиного эмбриона экспрессия sog ингибируется DL-зависимым репрессором sna . В то время как DPP присутствует во всей дорсо-латеральной зоне эмбриона, DPP-зависимый транскрипционный фактор MAD включается лишь вдоль узкой полоски наиболее дорсальных клеток-предшественников экстраэмбриональных мембран. Ограничение действия DPP вдоль дорсо-вентральной оси происходит за счет того, что белок Sog связывается с ним и блокирует его действие. Таким образом, умозрительно можно предсказать, что чем выше зона экспрессии sog , тем уже полоска pMAD (зона действия dpp).
Так почему же у комара две экстраэмбриональные мембраны, а у мухи – одна редуцированая? Ответ на этот вопрос дает анализ экспрессии генов на стадии бластодермы.
И у комара, и у мухи часть дорсальных генов (zen и hnt ) одинаково экспрессируются в дорсальном эпителии. А другие гены (tup и doc ) образуют петлеобразный рисунок, отличный от мушиного. Более того, гены hb и emc , традиционно детектируемые в головном отделе эмбриона мухи, экспрессируются в дорсальном эпителии комара как раз там, где наблюдается репрессия tup и doc .
Наконец, самым значительным отличием является то, что у комара sog экспрессируется в вентральных клетках эмбриона, а не в латеральной зоне, как у мухи. И зона спецификации экстраэмбриональных мембран у комара значительно шире, чем у мухи, так что в этой зоне умещаются два типа клеток вместо одного.
Суммируя все известное о процессе редукции двух типов ткани у комара к одному типу у мухи (рис. 6), можно предположить, что этот процесс является двухкомпонентным. Во-первых, наблюдается эволюция энхансера sog у мухи: дрозофильные сайты связывания dl лучше комариных, и вследствие этого уровень экспрессии sog у мухи «выше» комариного. Во-вторых, при увеличении зоны экспрессии sog уменьшается зона активности dpp , определяющего участок эмбриона, дифференцирующегося в экстраэмбриональную ткань. Зона активности dpp у мухи слишком мала для дифференциации двух типов тканей (как у комара). По-видимому, вследствие потери дорсальной экспрессии hb и emc происходит слияние двух типов клеток (как у комара) в один (у мухи). Таким образом, мы проследили эволюцию новой ткани от морфологии до генетики.
Куда же нам плыть?
Что возникает раньше – вопрос, на который хочется ответить, или же материал для исследований? Трудно представить себе первобытного человека, вышедшего из пещеры, взглянувшего на небо и немедленно задавшегося целью построить звездолет, да не простой, а чтобы непременно летал быстрее скорости света. Хотя бы по той причине, что во многих культурах звезды считались светильниками, закрепленными на голубой сфере, не говоря уже о представлениях о природе самого света. Таким образом, научный материал сам по мере изучения предоставляет ученому вопросы, и в этом плане роль науки весьма пассивна.
Тем не менее... Можно ли, сдвинув вентрально мушиный sog , воссоздать двухслойную структуру экстраэмбриональных мембран в мухе? Как работает энхансер sog у других насекомых? Можно ли, применив филогенетический метод, восстановить историю развития этого генетического элемента? Исходя из восстановленной истории можно ли составить некоторое представление об экологических силах, вызвавших эти генетические изменения?
Эти и многие другие вопросы... скорее всего, отпадут в ходе исследований за ненадобностью, поскольку они – всего лишь светильники на небосводе. Содержание этих вопросов вызвано не научным материалом, а лишь нашим представлением о нем. Так что будем двигаться дальше и посмотрим, какой сюрприз преподнесет нам природа, которую мы изучаем.
Список дополнительных и вспомогательных материалов
1. FlyMove – очень увлекательный сайт, где можно посмотреть ускоренное кино различных стадий развития мухи. http://flymove.unimuenster.de/Homepage.html
2. The Interactive Fly – более серьезный, образовательный сайт с описательным разделом по каждому изученному гену. http://www.sdbonline.org/fly/aimain/1aahome.htm
3. Еnsemble – интерактивная база данных, где хранятся отсеквенированные геномы. Можно без усилий добраться до нуклеотидной последовательности любого интересующего вас участка ДНК. http://www.ensembl.org/Drosophila-melanogaster/index.html
4. BDGP in situ – база данных, хранящая фотографии паттернов экспрессии генов мухи во время эмбриогенеза. Для примера впечатайте bcd и нажмите Enter. http://www.fruitfly.org/cgi-bin/ex/insitu.pl
5. FlyBase – научная база данных, содержащая информацию по существующим линиям мух. http://flybase.org/ PubMed – база данных по научной литературе. http://www.ncbi.nlm.nih.gov/entrez/query.fcgi
По материалам статьи Ю.Гольцева «Гены и Чебурашки: генетика возникновения новых форм»
Развитие насекомых
Индивидуальное развитие насекомых (онтогенез) складывается из эмбрионального развития, протекающего на фазе яйца, и постэмбрионального - после выхода личинки из яйца до достижения взрослой фазы - имаго.
Эмбриональное развитие . Яйца насекомых разнообразны по форме в связи с приспособлениями к среде, в которой они развиваются. Например, у жуков яйца преимущественно овальные и развиваются чаще в закрытом субстрате; у клопов - бочонковидные, прикрепленные к субстрату; у бабочек - башенковидные или бутылковидные; у сетчатокрылых (золотоглазок) яйца со стебельком. Часто яйца откладываются группами. Кладки яиц могут быть открытыми или закрытыми. Примером открытой кладки могут служить яйца колорадского жука, приклеенные самкой на нижнюю сторону листьев картофеля. К закрытым кладкам относятся кубышки саранчовых, образованные из частиц почвы, сцементированных выделениями придаточных желез самок. Тараканы откладывают яйца в оотеках - яйцевых капсулах, образующихся в половых путях самки.
Яйца насекомых снаружи покрыты оболочкой - хорионом, защищающим их от высыхания (рис. 337). На поверхности оболочки имеется микропиле - маленькое отверстие со сложной "пробкой" с канальцем внутри для проникновения сперматозоидов при оплодотворении. Под хорионом находится тонкая желточная оболочка, а под ней плотный слой цитоплазмы. Центральная часть цитоплазмы переполнена желтком. В цитоплазме расположены ядро и полярные тельца.
Дробление поверхностное. Вначале ядро многократно делится, дочерние ядра с участками цитоплазмы мигрируют к периферии яйца, покрываются мембраной и образуется поверхностный слой клеток - бластодерма, а в центре яйца остается желток. На брюшной поверхности бластодермы клетки более высокие и образуют утолщение - зародышевую полоску. Эта стадия эмбрионального развития насекомых соответствует бластуле.
Деление клеток зародышевой полоски ведет к развитию зародыша. Зародышевая полоска постепенно погружается, образуя брюшную борозду.
Рис. 337. Строение яйца насекомого (из Бей-Биенко): 1 - микропиле, 2 - хорион, 3 - желточная оболочка, 4 - ядро, 5 - полярные тельца, 6 - желток

Складки бластодермы над бороздой смыкаются, и образуются зародышевые оболочки: сероза и амнион (рис. 338). Здесь проявляются конвергенция с высшими позвоночными животными, у которых также имеются подобные оболочки. Благодаря образующейся амниотической полости зародыш оказывается во взвешенном состоянии внутри яйца, что надежно защищает его от механических повреждений Кроме того, жидкость, заполняющая амниотическую полость, облегчает обменные процессы зародыша.
Зародышевая полоска в дальнейшем дифференцируется на два слоя: нижний - эктодерму и верхний - энтомезодерму. Энтомезодерма у разных видов насекомых может образовываться по-разному: путем инвагинации или иммиграции клеток.
На следующем этапе развития эктодермальный слой полоски начинает по бокам загибаться вверх, а затем смыкается на спине, образуя замкнутую стенку зародыша. При смыкании стенок тела на спине часть желтка и желточных клеток попадает внутрь тела зародыша. Одновременно с образованием стенок тела зародыша в энтомезодерме обособляются две группы клеток на переднем и заднем концах тела. Это два зачатка средней кишки. В дальнейшем из этих двух зачатков начинают формироваться передний и задний участки средней кишки, которые потом смыкаются. Одновременно на переднем и заднем концах тела зародыша образуются глубокие впячивания эктодермы, из которых формируются передний и задний отделы кишки. Потом все три отдела соединяются, образуя сквозную кишечную трубку.
Мезодермальная полоска распадается на парные метамерные зачатки целомических мешков. Но в дальнейшем они распадаются, и из мезодермы образуются мускулатура зародыша, соматический листок целомического эпителия, сердце, жировое тело и гонады. Висцеральный листок целомического эпителия у насекомых не образуется, а полость тела становится смешанной - миксоцелью. Целомические зачатки сливаются с первичной полостью тела.
Позднее из эктодермы формируется нервная система и трахейная система. Из стенок задней кишки образуются мальпигиевы сосуды.
В процессе развития зародыш насекомого претерпевает сегментацию, которая вначале проявляется в передней части, а потом в задней части тела. В головном отделе закладываются акрон с глазными, губными и

антеннальными лопастями, интеркалярный сегмент и три челюстных сегмента. Затем оформляются три грудных и десять брюшных сегментов и анальная лопасть.
У многих насекомых зародыш проходит три стадии, характеризующиеся разным составом зачатков конечностей: протоподную, полиподную и олигоподную (рис. 339).
Для эмбрионального развития насекомых характерно явление бластокинеза. Это изменение положения тела зародыша в яйце, при котором наиболее полно используются запасы желтка.
Два типа бластокинеза для насекомых описаны А. Г. Шаровым. У насекомых с неполным превращением вначале зародыш располагается спиной вверх и головой к переднему концу яйца, а затем при образовании амниотической полости зародыш переворачивается брюшной стороной вверх, а голова соответственно оказывается в задней части яйца.
По-другому происходит бластокинез у большинства насекомых с полным превращением и у прямокрылых, у которых зародыш погружается в желток, не меняя положения тела в яйце.
В эмбриональном развитии насекомых проявляются приспособления к жизни на суше: защитные оболочки (хорион, сероза, амнион), запас питательных веществ (много желтка), амниотическая полость, заполненная жидкостью.
Перед вылуплением сформировавшаяся личинка насекомого заглатывает жидкость из амниотической полости, за счет чего усиливается тургор тела. Личинка прорывает хорион головой, на которой часто имеются яйцевые зубчики или шип.
Постэмбриональное развитие . В период постэмбрионального развития насекомых после выхода из яйца происходит рост молодого животного путем последовательных линек и прохождения качественно различных фаз развития. В течение онтогенеза, или индивидуального развития, насекомые линяют от 3-4 до 30 раз. В среднем число линек составляет 5-6. Промежуток между линьками называют стадией, а состояние развития - возрастом. Морфологические изменения в процессе развития от личинки до взрослого насекомого называются метаморфозом. У всех насекомых, кроме низших бескрылых форм, после достижения взрослого состояния - имаго рост и линьки прекращаются. Поэтому, например, вариации в размерах жуков одного вида нельзя относить к разным возрастным группам, а следует считать лишь проявлением индивидуальной изменчивости.
Различают три основных типа постэмбрионального развития насекомых: 1) прямое развитие без метаморфоза - аметаболия, или протометаболия; 2) развитие с неполным превращением, или с постепенным метаморфозом, - гемиметаболия; 3) развитие с полным превращением, т.е. с резко выраженным метаморфозом, - голометаболия.
Аметаболия , или прямое развитие, наблюдается только у первично-бескрылых насекомых из отряда щетинкохвостых (Thysanura), к которым относится часто встречающаяся чешуйница (Lepisma). Такой же тип развития наблюдается у Entognatha: коллембол (Collembola) и двухвосток (Diplura).
При аметаболии из яйца выходит личинка, похожая на имаго. Отличия касаются лишь размеров, пропорций тела и степени развития гонад. В отличие от крылатых насекомых, у них линьки продолжаются и в имагинальном состоянии.
Гемиметаболия - неполное превращение, или развитие с постепенным метаморфозом. Характерно для многих крылатых насекомых, например для тараканов, кузнечиков, саранчи, клопов, цикад и др.
При гемиметаболии из яйца выходит личинка, похожая на имаго, но с зачаточными крыльями и недоразвитыми гонадами. Такие имагоподобные личинки с зачатками крыльев называются нимфами. Это название заимствовано из древнегреческой мифологии и относится к божественным крылатым существам в облике девушек. Нимфы насекомых несколько раз линяют, и с каждой линькой зачатки крыльев у них увеличиваются. Нимфа старшего возраста линяет, и из нее выходит крылатое имаго. На рисунке 340 показаны фазы развития саранчи (яйцо, нимфы 1-5-го возрастов и имаго) как пример неполного превращения. Такое типичное неполное превращение называют гемиметаморфозом.
Среди насекомых с неполным превращением имеются случаи развития, когда нимфы заметно отличаются от имаго наличием особых личиночных приспособлений - провизорных органов. Такое развитие наблюдается у стрекоз, поденок, веснянок. Нимфы этих насекомых живут в воде, и их потому называют

Рис. 340. Развитие с неполным превращением у саранчи Locusta migratoria (по Холодковскому): 1 - переднефудь, 2 - среднегрудь с зачатками крыльев, 3 - заднегрудь с зачатками крыльев
наядами (водными нимфами). У них имеются такие провизорные органы, как трахейные жабры, исчезающие у сухопутных имаго. А у личинок стрекоз еще имеется "маска" - видоизмененная нижняя губа, служащая для схватывания добычи.
Голометаболия - полное превращение. Фазами развития при голометаболии являются: яйцо - личинка - куколка - имаго (рис. 341). Такое развитие характерно для жуков, бабочек, двукрылых, перепончатокрылых, ручейников и сетчатокрылых.

Рис. 341. Развитие с полным превращением у тутового шелкопряда Bombyx топ (по Лейнесу): А
- самец, Б - самка, В - гусеница, Г- кокон, Д - куколка из кокона
Личинки насекомых с полным превращением не похожи на имаго и часто отличаются экологически. Так, например, личинки майских жуков живут в почве, а имаго - на деревьях. Личинки многих мух развиваются в почве, гниющем субстрате, а имаго летают и посещают цветы, питаясь нектаром. Личинки таких насекомых несколько раз линяют и затем превращаются в куколку. На фазе куколки происходит гистолиз - разрушение личиночных органов и гистогенез - формирование организации взрослого насекомого. Из куколки выходит крылатое насекомое - имаго.
Таким образом, у насекомых наблюдаются следующие типы постэмбрионального развития: аметаболия, или протоморфоз (яйцо - личинка (похожая на имаго) - имаго); гемиметаболия - неполное превращение (яйцо - нимфа - имаго): гемиметаморфоз - типичный вариант, гипоморфоз - пониженный метаморфоз, гиперморфоз - повышенный метаморфоз; голометаболия - полное превращение (яйцо - личинка - куколка - имаго): голометаморфоз - типичный вариант, гиперметаморфоз - с несколькими типами личинок.
Типы личинок насекомых с полным превращением . Личинки голометаболических насекомых имеют более упрощенное строение по сравнению с имаго. У них нет сложных глаз, зачатков крыльев; ротовой аппарат грызущего типа, усики и ноги короткие. По развитости конечностей различают четыре типа личинок: протоподные, олигоподные, полиподные и аподные (рис. 342). Протоподные личинки характерны для пчел, ос. У них имеются лишь зачатки грудных ног. Эти личинки малоподвижны, развиваются в сотах с заботой со стороны рабочих особей. Олигоподные личинки встречаются чаще других, для них характерно нормальное развитие трех пар ходильных ног. К олигоподным относятся личинки жуков, сетчатокрылых. Полиподные личинки, или гусеницы, обладают, кроме трех пар грудных ног, еще несколькими парами ложных ножек на брюшке. Брюшные ножки представляют выступы брюшной
стенки тела и несут крючочки и шипики на подошве. Гусеницы характерны для бабочек и пилильщиков Аподные, или безногие, личинки наблюдаются в отряде двукрылых, а также у некоторых жуков (личинки усачей, златок), бабочек.

Рис. 342.Личинки насекомых с полным превращением (из Барнса): А - протоподная, Б, В - олигоподные, Г- полиподная, Д, Е, Ж
- аподные
По способам движения личинки насекомых с полным превращением подразделяются на камподеовидные с длинным, гибким телом, бегательными ногами и чувствующими церками, эруковидные с мясистым, слабоизогнутым телом с конечностями или без них, проволочниковые - с жестким телом, круглым в поперечнике, с опорными церками - урогомфами и червеобразные - безногие.
Камподеовидные личинки характерны для многих хищных жуков - жужелиц, стафилинов. Они передвигаются по скважинам в почве. Типичной эруковидной личинкой является личинка майского жука, навозников, бронзовок. Это роющие личинки. Проволочники характерны для жуков-щелкунов и чернотелок, личинки которых активно прокладывают ходы в почве. Червеобразных личинок множество. Они передвигаются в почве, тканях растений. К ним относятся не только личинки двукрылых, но и некоторых жуков, бабочек, пилильщиков, развивающихся, например, в тканях растений.
Типы куколок. Куколки бывают свободные, покрытые и скрытые (рис 343). У свободных куколок зачатки крыльев, конечностей хорошо видны и свободно отделены от тела, например у жуков. У покрытых куколок все зачатки плотно прирастают к телу, например у бабочек. Покровы у свободных куколок тонкие, мягкие, а у покрытых - сильно склеротизированные Еще выделяют тип скрытых куколок, покрытых затвердевшей несброшенной

Рис. 343. Типы куколок у насекомых (из Вебера): А - свободная жука, Б - покрытая бабочки, В - скрытая мухи; 1 - антенна, 2 - зачатки крыльев, 3 - нога, 4
- дыхальца
личиночной шкуркой, которая образует ложный кокон - пупарий. Внутри пупария находится открытая куколка. Поэтому скрытая куколка представляет собой лишь вариант свободной. Пупарий характерны для многих мух.
Нередко личинка последнего возраста перед окукливанием плетет кокон. Так, например, гусеница бабочек-шелкопрядов выделяет из шелкоотделительных желез шелк, из которого она свивает плотный кокон. Внутри такого кокона находится покрытая куколка. А у некоторых перепончатокрылых - муравьев, а также у сетчатокрылых внутри кокона находится открытая, или свободная, куколка. У личинок сетчатокрылых, например у золотоглазки, нити для кокона продуцируются мальпигиевыми сосудами и выделяются из анального отверстия.
Физиология метаморфоза . В процессе метаморфоза происходят два взаимосвязанных процесса: гистолиз и гистогенез. Гистолиз - это распад тканей личиночных органов, а гистогенез - образование органов взрослого насекомого. У насекомых с неполным превращением эти процессы протекают постепенно на фазе нимфы, а у насекомых с полным превращением - на фазе куколки.
Гистолиз происходит за счет деятельности фагоцитов и ферментов. При этом прежде всего разрушаются жировое тело, личиночные мышцы и некоторые другие органы, которые превращаются в питательный субстрат, расходуемый развивающимися тканями.
Гистогенез, или формирование органов взрослого насекомого, происходит главным образом за счет развития имагинальных дисков - зачатков из недифференцированных клеток. Имагинальные диски закладываются еще на личиночной фазе и даже в эмбриогенезе и представляют собой внутренние зачатки. Из имагинальных дисков развиваются глаза, крылья, ротовой аппарат, ноги, а также внутренние органы: мускулатура, гонады. Пищеварительная система, мальпигиевы сосуды, трахеи не разрушаются, а сильно дифференцируются в процессе метаморфоза. Менее всего метаморфизируется сердце и нервная система. Однако при метаморфозе в нервной системе часто наблюдается процесс олигомеризации (слияния) ганглиев.
Процесс метаморфоза контролируется железами внутренней секреции (рис. 329). Нейросекреторные клетки мозга выделяют гормоны, активизирующие деятельность кардиалъных тел , гормоны которых через гемолимфу стимулируют проторакалъные (переднегрудные) железы, выделяющие линочный гормон - экдизон . Экдизон способствует процессу линьки: частичному растворению и отслаиванию старой кутикулы, а также формированию новой.
В процессе метаморфоза существенное значение имеет также деятельность прилежащих тел , продуцирующих ювенильный гормон. При высокой его концентрации линька личинки приводит к образованию личинки
следующего возраста. По мере роста личинок деятельность прилежащих тел ослабевает и падает концентрация ювенильного гормона, а переднегрудные железы постепенно дегенерируют. Это приводит к тому, что личинки линяют в фазу куколки, а затем имаго.
Искусственная пересадка прилежащих тел, например в нимфу саранчовых последнего возраста, способствует тому, что она линяет не во взрослую фазу, а в более крупную личинку дополнительного возраста. На фазе имаго ювенильный гормон контролирует развитие гонад, а гормон экдизон уже не вырабатывается в связи с редукцией переднегрудных желез.
Происхождение метаморфоза . Существует несколько гипотез о происхождении метаморфоза у насекомых. Долгое время велись споры, какие насекомые в большей мере эволюционно продвинуты - с полным или неполным превращением. С одной стороны, нимфы насекомых с неполным превращением более прогрессивно развиты, чем личинки насекомых с полным превращением, с другой стороны, у последних имеется продвинутая фаза куколки.
В настоящее время это противоречие снято гипотезой о происхождении метаморфоза Г. С. Гилярова, А. А. Захваткина и А. Г. Шарова. Согласно этой гипотезе, обе формы метаморфоза у насекомых развивались независимо от более простого типа развития - протоморфоза, наблюдаемого у первичнобескрылых насекомых, например у щетинкохвостых (Thysanura).
При протоморфозе развитие прямое, при этом наблюдается множество линек на фазе личинок, а затем и в имагинальном состоянии. Все фазы развития этих насекомых протекают в одной и той же среде.
Предполагается, что в процессе эволюции насекомые переходили от полускрытого существования в верхнем слое почвы к обитанию на ее поверхности и на растениях. Этот переход в новую среду обитания увенчался крупным ароморфозом - развитием крыльев и полета.
Освоение открытых местообитаний отразилось на эволюции индивидуального развития насекомых. Эволюция онтогенеза насекомых, по-видимому, шла по двум основным направлениям.
В одном случае шел процесс эмбрионизации развития, приводивший к вылуплению насекомых из яиц, богатых желтком, на более поздних фазах развития. Это привело к имагинизации личинок с образованием нимф. Так развились насекомые с неполным превращением. Этот путь эволюции привел к прогрессивному развитию личинок, ведущих сходный образ жизни с имаго.
В другом случае, наоборот, происходил процесс дезэмбрионизации развития, т. е. осуществлялся выход из яиц, бедных желтком, на более ранних фазах развития. Это привело к морфоэкологическому расхождению личинок и имаго насекомых. Личинки упростились и приспособились
к обитанию в более защищенной среде, выполняя функцию питания, а имаго стали в основном выполнять функцию размножения и расселения. Кроме дезэмбрионизации развития личинок насекомых с полным превращением, у них развилось множество провизорных адаптации к различным условиям существования. Таким образом, произошел амфигенез (расхождение) в эволюции личинок и имаго у насекомых с полным превращением. Амфигенез личинок и имаго оказался очень глубоким по морфологическим адаптациям, что создавало серьезные противоречия в онтогенезе. Они благополучно были разрешены путем возникновения фазы куколки, во время которой происходит радикальная перестройка личиночной организации на имагинальную. Это позволило насекомым с полным превращением освоить более широкий спектр экологических ниш и достичь небывалого расцвета среди животных на Земле.
Размножение насекомых . Для большинства насекомых характерно обоеполое половое размножение. У многих видов выражен половой диморфизм. Например, у самцов жуков-оленей мандибулы видоизменены в рога, а у самцов жуков-носорогов имеются рог на голове и горбы на переднеспинке. Это связано с брачным поведением этих видов, сопровождающимся борьбой самцов за самку. Взаимоотношения между полами у разных видов чрезвычайно разнообразны. Самцы двукрылых семейства Dolichopodidae приносят самке "подарок" - пойманную мушку и исполняют танец с зеркальцами на ножках. Самки богомолов отличаются хищным нравом и съедают самца во время спаривания.
Большинство насекомых откладывают яйца, но нередко наблюдается и живорождение. В этом случае яйца развиваются в половых путях самки и она рождает личинок. Так, мясные мухи-саркофаги (Sarcophagidae) откладывают на мясо живых личинок, развитие которых протекает очень быстро. Недаром в древности считали, что черви в мясе самозарождаются. Можно не заметить, как мясная муха посетила открыто лежащее мясо, и неожиданно обнаружить внезапно появившихся белых личинок.
К живородящим видам относятся также муха овечья кровососка, некоторые жуки, обитающие в пещерах.
Кроме обоеполого полового размножения у ряда насекомых наблюдается партеногенез - развитие без оплодотворения. Имеется немало видов из разных отрядов насекомых, для которых характерен партеногенез. Партеногенез может быть облигатным - обязательным, тогда все особи вида только самки. Так, в высокогорных условиях, на севере и в других неблагоприятных условиях встречаются партеногенетические жуки, прямокрылые, уховертки, сетчатокрылые. Партеногенез встречается и у обоеполых видов, когда часть яиц откладывается оплодотворенными, а- часть без оплодотворения. Например, трутни у пчел развиваются из неоплодотворенных яиц.
Подобный партеногенез встречается и у других перепончатокрылых (муравьев, пилильщиков), у термитов, некоторых клопов и жуков. А у тлей, например, происходит смена поколений в жизненном цикле: обоеполого и партеногенетического. В некоторых случаях партеногенез может быть факультативным (временным), проявляющимся только при неблагоприятных условиях. Партеногенез у насекомых способствует поддержанию высокой численности популяций.
Вариантом партеногенеза является педогенез - размножение без оплодотворения на личиночной фазе развития. Это особый путь развития насекомых, когда созревание гонад опережает другие органы. Так, например, некоторые виды комариков-галлиц размножаются на личиночной фазе. Личинки старших возрастов рождают личинок младших возрастов. Педогенез отмечен для одного из видов жуков, личинки которого частично откладывают яйца, частично рождают личинок. Педогенез в дополнение к обоеполому размножению увеличивает численность вида.
Жизненные циклы насекомых . В отличие от онтогенеза, или индивидуального развития насекомых, жизненный цикл - это развитие вида, включающее, как правило, несколько типов онтогенезов. Онтогенез ограничивается жизнью одной особи от яйца до наступления полового созревания и затем естественного отмирания. Жизненный цикл - это повторяющаяся часть непрерывного процесса развития вида. Так, в наиболее типичном случае у насекомых жизненный цикл складывается из двух сопряженных и отличающихся между собой морфофизиологически онтогенезов самцов и самок, размножающихся половым путем и воспроизводящих себе подобных. А у партеногенетических видов жизненный цикл характеризуется лишь одним типом онтогенеза самок.
Жизненные циклы насекомых разнообразны по типам размножения, составу поколений и их чередованию. Можно выделить следующие типы жизненных циклов насекомых.
1. Жизненные циклы без чередования поколений с обоеполым половым размножением. Это наиболее распространенный тип жизненного цикла, характерный для диморфных видов, состоящих только из самцов и самок, размножающихся половым путем. Таковы циклы у большинства жуков, бабочек, клопов.
2. Жизненные циклы без чередования поколений с партеногенетическим размножением. Такие виды мономорфные, состоящие только из партеногенетических самок, откладывающих яйца без оплодотворения.
Партеногенетические виды особенно часто встречаются среди тлей, листоблошек и других равнокрылых. В высокогорных условиях распространены Партеногенетические виды жуков, клопов, кузнечиков, кокцид.
3. Редчайшим типом жизненного цикла у насекомых является цикл без чередования поколений с половым размножением гермафродитных видов.
Известен американский вид мухи, состоящий только из гермафродитных особей. На ранних фазах развития имаго функционируют как самцы, а на поздних - как самки. Поэтому все особи откладывают яйца, что повышает численность вида.
4. Жизненные циклы без чередования поколения с половым размножением и факультативным партеногенезом у полиморфных видов, например у общественных насекомых. Вид состоит из половых особей - самцов и самок и фертильных - рабочих особей, не участвующих в размножении. К таким видам относятся пчелы, муравьи, термиты. Такие жизненные циклы осложнены тем, что самки откладывают наряду с оплодотворенными яйцами и Партеногенетические, из которых, например, у пчелиных развиваются гаплоидные самцы - трутни, а из оплодотворенных - самки и рабочие особи женского пола. Подобно развиваются некоторые наездники, трипсы, кокциды.
У других видов факультативный партеногенез проявляется по-другому: из неоплодотворенных яиц развиваются не самцы, а самки. Но в этом случае у самок восстанавливается диплоидный набор хромосом путем слияния гаплоидных ядер. Такое развитие известно у некоторых палочников, саранчовых, пилильщиков, кокцид.
5. Жизненные циклы с чередованием полового поколения и партеногенетического (гетерогония). У многих тлей и филлоксеры, кроме полового поколения крылатых самцов и самок, имеется несколько сменяющихся поколений партеногенетических самок, крылатых или бескрылых.
6. Жизненные циклы с чередованием полового поколения и нескольких поколений с педогенезом. Например, у некоторых комариков-галлиц после полового размножения, в котором участвуют самцы и самки, происходит партеногенетическое размножение личинок (педогенез). После нескольких поколений размножающихся личинок, отмирающих после живорождения себе подобных, последнее поколение личинок окукливается и дает крылатых самок и самцов.
7. Жизненные циклы с чередованием полового поколения (самцов и самок) с бесполым. После полового размножения самки откладывают оплодотворенные яйца, которые претерпевают полиэмбрионию. Это бесполое размножение на фазе зародыша. Яйцо претерпевает дробление, и зародыш на фазе морулы начинает размножаться путем почкования. Из одного яйца может образоваться несколько десятков зародышей. Такие
Таким образом, классификацию жизненных циклов можно представить следующим образом.
I. Без чередования поколений :
- 1) с обоеполым половым размножением (майский жук);
- 2) с партеногенетическим размножением (высокогорные жуки, кузнечики);
- 3) с половым размножением гермафродитных особей (американская муха);
- 4) с половым размножением и частичным партеногенезом у полиморфных видов (пчелы).
II. С чередованием поколений :
- 1) гетерогония: чередование полового поколения и нескольких партеногенетических (тли, филлоксера);
- 2) гетерогония: чередование полового поколения и нескольких педогенетических поколений (некоторые галлицы);
- 3) метагенез: чередование полового поколения с полиэмбрионией (наездники).
Сезонные циклы насекомых . Если под жизненным циклом понимается циклически повторяющаяся часть морфогенеза вида от одной фазы развития до одноименной, то под сезонным циклом развития понимается характеристика развития вида в течение сезонов одного года (от зимы до зимы).
Так, например, жизненный цикл майского жука протекает в течение 4-5 лет (от яйца до половозрелых имаго), а для сезонного цикла этого вида характерно, что весной перезимовавшие личинки окукливаются и молодые жуки размножаются. Летом, осенью и зимой встречаются их личинки разных возрастов. Число поколений, развивающихся в течение года, называется вольтинностью.
Различаются виды, дающие несколько поколений в год. Это поливольтинные виды. Например, комнатная муха может давать 2-3 поколения за сезон и зимует в фазе имаго. Большинство насекомых моновольтинные, дающие одно поколение за год.
Сезонные циклы насекомых в природе характеризуются календарными сроками встречаемости различных фаз развития. Важными особенностями сезонных циклов видов являются сроки их активной жизни и диапаузы (временной задержки развития) зимней или летней. Регуляцию жизненных циклов видов в соответствии с местными сезонными явлениями обеспечивают факторы среды и нейрогуморальная система организма.
Лесная энтомология*
Развитие насекомых
Развитие насекомых делится на два периода: зародышевое , или эмбриональное (внутри яйца), и внеяйцевое , или постэмбриональное .
Зародышевое развитие начинается в яйце дроблением ядра с момента оплодотворения или откладки яиц. Яйцо имеет круглую, овальную, удлиненную или другую форму и представляет собой одну крупную клетку, включающую питательный желток, необходимый для роста и развития зародыша. Снаружи яйцо покрыто оболочкой (хорион ), играющей роль скорлупы. На одном из полюсов яйца имеется одно или несколько мельчайших отверстий (микропиле ), через которые проникают сперматозоиды при оплодотворении. Яйцо после откладки часто всасывает влагу из окружающей среды и вследствие этого увеличивается в 2-3 раза.
Зрелые яйца самки откладывают поодиночке или группами на листья, ветви и стволы деревьев, на почву, травянистую растительность и другие предметы. Часто яйца бывают погружены в субстрат (в древесную ткань или почву) или защищены пушком, снятым с конца брюшка самки (у златогузки, непарного шелкопряда и других бабочек), или щитком, образовавшимся из выделений придаточных желез (у зеленой узкотелой златки).
Развитие зародыша в яйце продолжается от нескольких дней до месяца и более. Оно заключается в ряде сложных преобразо.ваний и заканчивается вылуплением личинки, прогрызающей оболочку яйца:
Вылупление личинки из яйца звездчатого ткача
Постэмбриональное развитие начинается после вылупления из яйца личинки и продолжается от нескольких дней до нескольких лет. Рост личинки сопровождается линьками , которые стимулируются деятельностью желез внутренней секреции. При этом в кровь поступает гормон, вызывающий деление клеток гиподермы и выделение ими новой кутикулы. В то же время кожные личиночные железы начинают изливать под старую кутикулу секрет, который растворяет внутреннюю кутикулу снизу, отчего она лопается по средней линии спины и сбрасывается.
Линьками, разделены возрасты личинок. Число возрастов различно. У многих личинок жуков всего три, у гусениц бабочек чаще всего пять, а у поденок до 45. Количество возрастов может меняться в зависимости от температуры и пищи при голодании.
В процессе постэмбрионального развития насекомые не только растут, но и претерпевают ряд изменений. Совокупность этих изменений называется превращением , или метаморфозом . Основной момент метаморфоза состоит в приобретении крыльев. В зависимости от этого различают два основных типа метаморфоза: неполный (насекомые с неполным превращением - прямокрылые, клопы, цикады, тли, стрекозы и др.) и полный (насекомые с полным превращением - жуки, бабочки, перепончатокрылые, мухи).
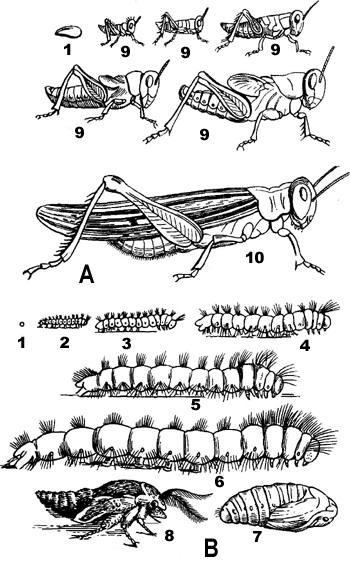
Метаморфоз у насекомых : А - неполное превращение (разные возрасты саранчи); В - полное превращение соснового шелкопряда. 1 - яйцо, 2-6 - гусеницы в различных возрастах, 7 - куколка, 8 - только что вылупившаяся бабочка, 9-10 - личинки и взрослые насекомые
Насекомые с неполным превращением характеризуются наличием трех фаз развития: яйца , личинки и взрослого насекомого (имаго). Выходящая из яйца личинка у них близка по внешнему виду и биологически к взрослому насекомому: туловище расчленено на грудь и брюшко, которые несут соответствующие придатки. Крылья появляются в виде наружной складки кожи на краю грудного сегмента. Она увеличивается при каждой линьке и постепенно достигает окончательной величины и формы. Пищевой режим у личинки такой же, как у взрослого насекомого.
Насекомые с полным превращением имеют четыре фазы развития: яйцо, личинку, куколку и взрослое насекомое (имаго). Выходящая из яйца личинка червеобразной формы. Ее туловище состоит из серии одинаковых сегментов. Брюшко и грудь личинки не обособлены; имеются только сегменты, из которых они потом образуются. Таким образом, личинка резко отличается от взрослого насекомого своим внешним видом. Личинка питается чаще всего совершенно по-другому и ведет образ жизни иной, нежели взрослое насекомое. После последней линьки личинка превращается в куколку , по своему внешнему виду похожую на взрослое насекомое, иногда покрытую оболочкой. Перед превращением в куколку личинка часто покрывает себя коконом, материал для которого готовят шелкоотделительные железы (бабочки), а иногда выделяют мальпигиевы сосуды (некоторые жуки, сетчатокрылые).
При полном превращении происходит сложная перестройка организма, начинающаяся у личинки перед окукливанием и заканчивающаяся у куколки. На протяжении этой перестройки временные органы личинки заменяются органами, характерными для взрослого насекомого. Зачатки органов взрослого насекомого скрыты у личинки в разных частях тела и называются имагинальными дисками . Из них развиваются крылья, ноги и ротовые органы взрослого насекомого, а также трахеи, мускулатура, пищеварительная система и др.
Перестройка слагается из двух процессов: гистолиза и гистогенеза . Гистолиз состоит в разрушении тех органов, которые функционируют у личинки и отсутствуют у взрослого насекомого. Разрушение происходит благодаря оживившейся деятельности кровяных клеток, которые активно внедряются в органы и поглощают куски ткани. Существует мнение, что этот процесс происходит химически, а кровяные клетки только подхватывают продукты распада. Гистогенез представляет образование различных тканей из недифференцированного клеточного материала - продуктов гистолиза. Активным элементом в гистогенезе служат имагинальные диски. Количество имагинальных дисков зависит от интенсивности метаморфоза и особенно велико у мух.
Весь процесс метаморфоза тесно связан с работой желез внутренней секреции . Гормон экдизон регулирует нормальное развитие личинки, способствует развитию у нее гонад, дифференциации тканей и линьке. Ювенальный гормон препятствует линьке во взрослую фазу и стимулирует рост и развитие личиночных органов.
У насекомых встречается ряд видоизменений неполного и полного метаморфоза. Так, при неполном превращении иногда наблюдается почти полное отсутствие изменений у личинки в процессе ее жизни или, наоборот, приближение к полному метаморфозу (гиперморфоз ). В этом случае у взрослой личинки приобретается неподвижность, и она получает название нимфы , напоминая куколку при полном превращении. Однако она не утрачивает прежнего сходства с личинкой.
При полном превращении наблюдается иногда несколько форм личинок, последовательно сменяющих друг друга в процессе роста и развития. Это связано с резкими изменениями их образа жизни в течение развития. Примером могут служить личинки нарывников. Сначала личинка очень неподвижна. Она проникает в кубышки саранчовых и гнезда, где питается яйцами диких пчел, а затем покидает их и проходит ряд последующих фаз. Такое превращение называется гиперметаморфозом .
* При составлении данного справочника использовалась книга А.И.Воронцова "Лесная энтомология. Учебник для вузов" (Изд. 3-е, перераб. М., "Высш. школа", 1975. 368 стр.) с изменениями.
Размножение и развитие. Насекомые размножаются половым путем.Различают два вида полового размножения-когда яйца оплодотворяются сперматозоидами (мужскими клетками) и когда зародыш развивается из неоплодотворенного яйца.Это девственный способ размножения, или партеногенез.Весь цикл развития насекомых разделяется на эмбриональное (внутри яйца) развитие и постэмбриональное.Развитие зародыша в яйце продолжается от нескольких дней до одного месяца.Постэмбриональное развитие начинается после вылупления личинки и продолжается от нескольких дней до нескольких лет.Личинки в процессе своего развития питаются и растут,причем рост сопровождается линьками,когда сбрасывается старая кутикула.Количество линек у различных насекомых различно.В процессе своего развития насекомые претерпевают ряд изменений.По характеру и особенностям метаморфоза насекомые разделяются на две группы:насекомые с неполным превращением и насекомые с полным превращением.Насекомые с неполным превращением имеют три фазы развития:яйцо,личинку и взрослое насекомое.Причем личинка очень похожа на взрослое насекомое.Насекомые с полным превращением имеют четыре фазы развития:яйцо,личинку,куколку,взрослое насекомое.Здесь личинки резко отличаются от взрослого насекомого (например,бабочка и ее личинка-гусеница).Личинка линяет несколько раз. С последней линькой она превращается в куколку-стадию покоя.Через некоторое время из куколки выходит взрослое насекомое,которое некоторое время остается неподвижным.За это время нежный кожный покров насекомого постепенно затвердевает и окрашивается, крылья расправляются.Насекомое во взрослой фазе неспособно к росту и не линяет.Некоторые насекомые после превращения во взрослую фазу имеют вполне развитые половые органы.Эти насекомые не питаются и живут очень недолго.Например,точильщики и моли.Период развития насекомых от яйца до половозрелого состояния называется жизненным циклом,генерацией или поколением.Продолжительность генерации у различных насекомых различна.Например,ветчинный кожеед имеет одно поколение в году,а хлебный точильщик при определенных условиях может иметь до четырех поколений в год.